Microtubule networks
Similar to the formation of actin filaments from the self-assembly of actin monomers, tubulin forms asymmetric dimers that can self-assemble into microtubules. Compared with actin filaments, microtubules are much more rigid, and almost straight in the dimensions of a single cell. Microtubules can sustain much higher
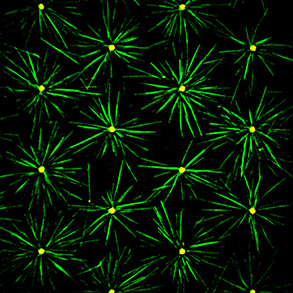
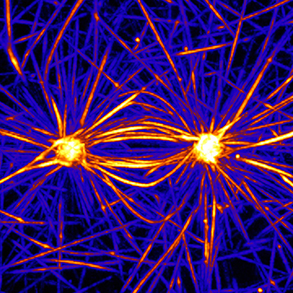
compression forces than actin filament but are not as numerous as actin filaments. In most animal cells, the MT network forms as an aster in which microtubules radiate from the microtubule organizing center (MTOC). As cells divide, the MTOC is duplicated and the network forms a bipolar spindle. Our aim is to investigate the
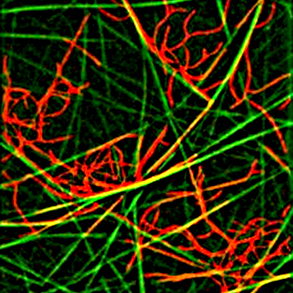
role of microtubule mechanics and microtubule-associated proteins in the establishment of the various microtubule network architectures during interphase and mitosis.